
大分県の植生
1.大分県の植生を支配する環境要素
大分県は九州本島の北東部、北緯32゜43’~33゜44’、東経130゜50’~132゜11’に位置しており、県東部は黒潮が洗う外洋に面し、県北部は内海の周防灘に面している。内陸地は県の北西部から西部にかけて英彦山や津江山地、南西部は九重火山群、南部は祖母・傾山系の峻嶺が屏風のように連なって、大分県を半円状に囲んでいる。
本県の気候は大局的には暖温帯夏雨型多雨気候域に属しており、県南部は夏の降水量が多い典型的な西南日本太平洋型気候域であるが、県北部は年間降水量が1600mm以下となって瀬戸内型気候へ移行する。また県北西部の山地は冬季北西季節風をまともに受けて、日本海型気候の影響が植生にも反映されている。沿岸部の植生は海流の影響が大きく、黒潮が洗う県南部の沿岸地域は、緯度に比べてより暖かく、暖地性の植物を多く伴った植生が成立する。大分県の気候区を川西博(1988)は瀬戸内型Ⅰ、瀬戸内型Ⅱ(南海型から瀬戸内型への移行帯)、南海型、内陸山地型の4つに分けているが、この気候区分と大分県の植生の地域性はよく一致している。
地質と植生の関係についてみると、西南日本の地質構造を内帯と外帯に2分する中央構造線が、大分県の中央を東西に走っており、これを境にして対照的な植生が成立する。
外帯の県南部は新生代第三紀に形成された祖母・傾山系のような古い火山が一部にあるが、大部分の地域は古生代や中世代に形成された三波川帯、秩父帯、四万十帯などの基盤岩が露出する非火山地帯であって、ここにはソハヤキ要素といわれる西南日本外帯に固有の植物が生育している。また壮年期に達した祖母・傾山系の地形は険しく、ここには気候を反映して、常緑広葉樹林帯とブナ帯の間にモミ・ツガ帯が成立する。
内帯の県北部や中部は新生代第四紀の火山活動による火山噴出物によって広く覆われている。県中部は九重火山群や由布・鶴見火山群などの、なだらかな裾野が広がり火入れや放牧が行われて、県南部の祖母・傾山系とは対照的な景観となる。フロラは氷期の頃に南下した遺存植物が多くみられ、火山山頂帯植生などの特有な植生が成立する。
2.大分県の植生の地域性
大分県の植生は太平洋型気候、瀬戸内型気候、内陸山地型気候といった気候的要因、内帯と外帯、火山地帯と非火山地帯といった土地的要因、これに地史や海流などの要因が重なって、地域に特有の植生が成立する。この植生の地域性は図1のように6つに大別できる。
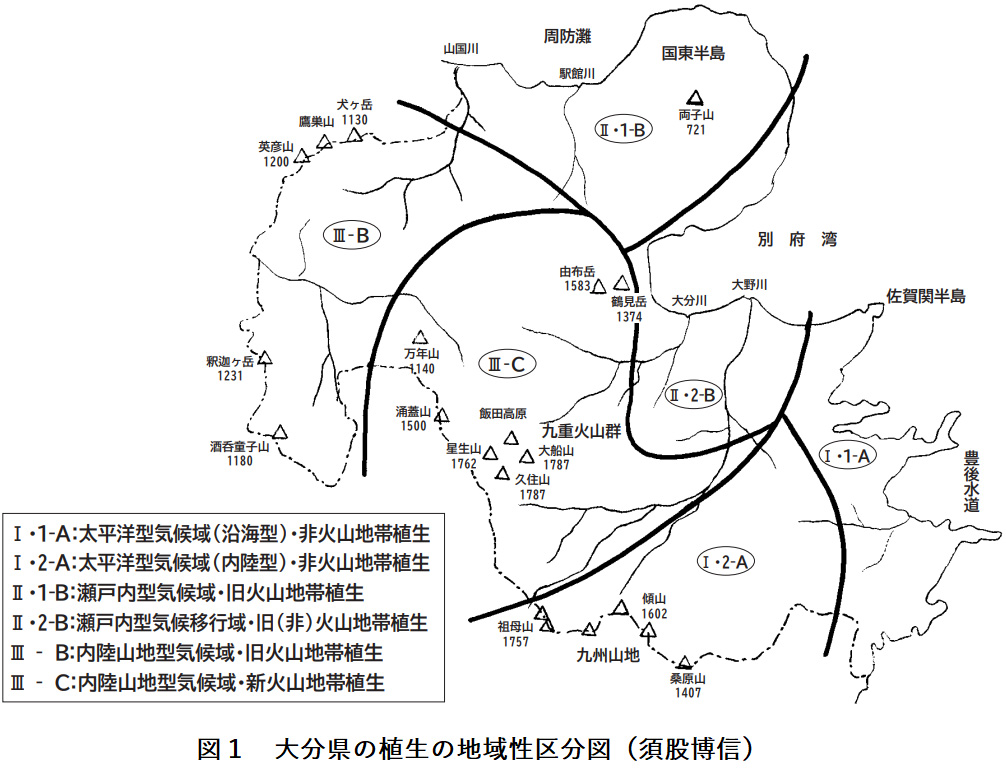
Ⅰ・1-A:太平洋型気候域(沿海型)・非火山地帯の植生
太平洋型気候域・非火山地帯の植生は東部の沿海地域と西部の内陸地域で大きな地域差が認められ、沿海型と内陸型に分かれる。これは高度的な差違ばかりでなく黒潮効果が深く関わっている。
この地域は九州山地の東端が豊後水道に沈んでリアス海岸を形成し、これを黒潮が洗って沿岸部は無霜地帯となる。豊後水道に突出した半島の崖地は、地史や土地的な要因を反映して、常緑硬葉型のウバメガシ-トベラ群集が成立する。また潮風が強い沿岸にはマサキ-トベラ群集、ハマビワ-オニヤブソテツ群集が成立し、ここには黒潮効果を反映してアコウ、ビロウ、ハマオモトなどの暖地性植物が自生する。年平均気温17℃、年間降水量2000mmを超えるこの地域は、沿岸部にタブ-ホソバカナワラビ群集やスダジイ-タイミンタチバナ群集、やや内陸部へ入るとコジイ-クロバイ群集、イチイガシ群集、ウラジロガシ-サカキ群集、アカガシ-ミヤマシキミ群集などが成立する。なお低地の神社林にはハナガガシの残存林があり、この群落の北限地になっている。また尾根の岩角地には九州東部の外帯に特有のアカマツ-オンツツジ群集が成立する。
Ⅰ・2-A:太平洋型気候域(内陸型)非火山地帯の植生
この地域は祖母山(1756.4m)、傾山、新百姓山、夏木山、桑原山など、標高1400~1700mの峻峰が続く九州山地とその山麓の地域である。古生代の基盤岩に第三紀の祖母火山岩が貫入してできたこの山系は、ソハヤキ要素といわれる西南日本外帯に固有の植物が生育しており、太平洋型気候を反映して図2の概念図のような森林帯が発達している。
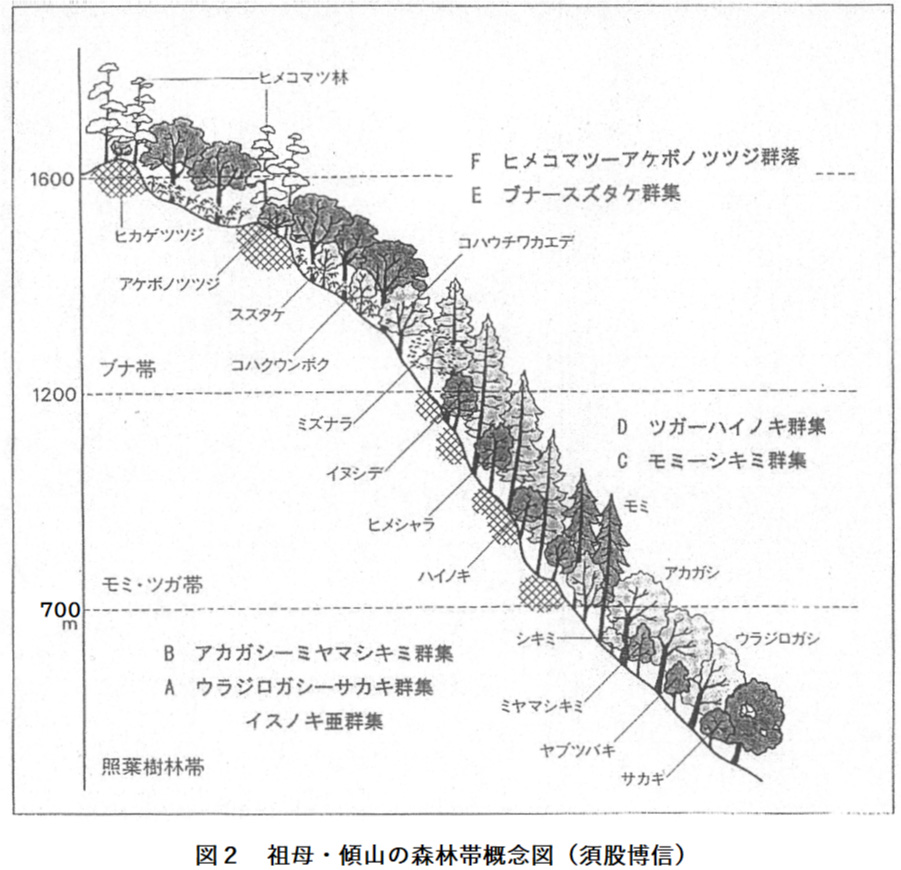
植物の活動期間の尺度を月平均気温10℃以上の月を生育期間とし、それ以外は休眠期間としてあらわすと、大分県は標高400mまでは生育期間8か月であって、シイやイチイガシの群集が成立する。標高500mで生育期間は7か月となるが空中湿度が高くなってウラジロガシ-サカキ群集やアカガシ-ミヤマシキミ群集が成立する。標高700mで暖かさの指数85、寒さの指数-9.1になって、この付近が常緑広葉樹林帯の上限になる。
標高700mから1200mの間は生育期間6か月であって、西南日本外帯に特有のモミ-シキミ群集、ツガ-ハイノキ群集が成立する。これらの群集はヤブツバキクラスの標徴種を伴った針葉樹林で中間温帯の性格が強い。
標高1200mを越えて生育期間5か月になるとブナ-スズタケ群集が成立し、暖かさの指数60、寒さの指数-20付近がツガ帯とブナ帯の境界になっている。
日本の亜高山帯針葉樹林は暖かさの指数15~45、生育期間1か月以上~4か月未満の高度に成立している。大分県は標高1700mで暖かの指数43であるが、標高1800mになっても生育期間は4か月であって、なおブナ林の成立を許す温度条件である。そのため大分県の森林帯はブナ帯までであって亜高山帯針葉樹林は成立しない。
壮年期に達した祖母・傾山系の地形は険しく、尾根の岩角地はヒメコマツ-ヒカゲツツジ群集が成立し、深い渓谷はシオジ-ミヤマクマワラビ群集が成立する。
Ⅱ・1-B:瀬戸内型気候域・旧火山地帯の植生
周防灘に面した地域は年平均気温15℃、年間降水量1500~1600mmであって夏季の雨量が少ない地域である。また冬季は周防灘を吹きぬける冬季北西季節風の影響でしばしば降雪をみる。この気候を反映して、沿岸地域のシイ林はスダジイ-ヤブコウジ群集、内陸部はコジイ-クロキ群集が成立する。また、凝灰岩上は夏季の乾燥も加わってイブキシモツケ-イワヒバ群集、イワシデ群落、アカマツ-ヤマツツジ群集が成立する。
Ⅱ・2-B:瀬戸内型気候移行域・旧(非)火山地帯の植生
別府湾に面した地域で太平洋型気候から瀬戸内型気候への移行帯に相当する。アコウ、ハマオモトなどの暖地植物やウバメガシ-トベラ群集の生育地は、佐賀関半島の南側までであって北側には及ばない。豊後水道を北上した黒潮が国東半島の南東部沿岸にぶつかる付近は、局所的にスダジイ-タイミンタチバナ群集が成立するが、そのほかの沿岸地域はスダジイ-ヤブコウジ群集のイズセンリョウ亜群集である。またⅡとⅢの気候域はタブ-イノデ群集が成立する。
Ⅲ-B:内陸山地型気候域・旧火山地帯の植生
第三紀中新世から第四紀更新世の火山岩類からなる英彦山、釈迦ヶ岳、酒呑童子山など標高1100~1200mの山々が連なる古い火山地帯である。年間降水量は2000~2600mm、標高80mの日田盆地は年平均気温14.3℃、水系に恵まれて、しかも気温の日較差が大きいために霧が発生しやすい。このような気象条件であるために林業が盛んで、丘陵帯から低山地帯はスギ植林地に開発されている。
古い火山地帯であるため、祖母・傾山系と同様に気候を反映した森林帯が発達している。しかし、英彦山のブナ林はスズタケのほかにクマイザサを伴って、太平洋型気候要素の退行が目立つようになる。丘陵帯では中国地方西部や九州東北部に特有のシイモチ-シリブカガシ群集(コジイ-クロキ群集)が成立し、低山地帯のツガ林では太平洋側要素のヒメシャラがヒコサンヒメシャラに代り、日本海側要素タムシバを伴うなど、日本海型気候の影響が強まってくる。
また犬ヶ岳や鷹ノ巣山などの筑紫溶岩上には第三紀遺存植物群落のヒノキ-ツクシシャクナゲ群集が残存し、耶馬溪溶結凝灰岩上にはイワシデ群落が成立する。
Ⅲ-C:内陸山地型気候域・新火山地帯の植生
九州本島最高峰の中岳(1791m)や大船山、久住山、稲星山、星生山、三俣山など、標高1700m級のトロイデ火山が集まった九重火山群、並びに由布・鶴見火山群とその山麓地域であって、今も一部で硫気が噴出するなど、地質年代の若い火山地域である。標高828mの飯田高原は年平均気温11.3℃、冬季の積雪日数は年平均30日、時には30~50㎝の積雪を見ることもある。
九重火山群は祖母・傾山系と緯度や海抜高度がほぼ同じであるが、土地的な要因が異なるために、祖母・傾山系のような森林帯はできないまま、火山地帯特有の植生が成立する。
山頂付近の植生は火山活動や冬季北西季節風の影響などもあって、標高1500m付近が火山森林限界になる。山頂部は高山植物のコケモモを伴ってミヤマキリシマ-マイヅルソウ群集が成立し、高山帯の風衝ハイデの相観によく似た火山山頂帯植生が成立する。
山地帯ではブナ-スズタケ群集が九重火山群においてもみられるが、この地域ではミヤコザサが優勢であって、ミズナラ林と結びつく。また野火の影響もあって、ノリウツギ-ヤマカモジ群集、ミズナラ-リョウブ群集が成立する。九重火山群の黒岳では火山噴出岩の影響で、ブナとササの結びつきが離れ、林冠群はコハウチワカエデが優占種のブナ-ツクシシャクナゲ群集が成立する。
低山地帯のモミ-ツガ林は火山ガスに弱いようであって、残存林は少ないが九重火山群の北山麓・鳴子川渓谷の断崖斜面に、小規模であるが残存林がある。九重火山群や由布・鶴見火山群の低地帯では、人為が加わってクヌギ-ミツバツチグリ群集、コナラ-コバノガマズミ群集、クマシデ-コガクウツギ群集などが成立する。
山麓の火山高原一帯は定期的な火入れ、採草、放牧などの人為が加わってススキ-トダシバ群集が広範囲にわたって成立する。また局所的ではあるが、凹地形の湿地にはヨシ-ヤマアゼスゲ群集やヌマガヤ-ヒメミズゴケ群集で代表される湿原植物群落を形成する。
丘陵地帯は河川に沿って火山溶岩や火砕流が流入している。大部分が安山岩類であって、ここにはシイ群落が成立しにくく、代ってアラカシ-ジャノヒゲ群集が成立する。
3.植生の垂直分布
祖母・傾山系と九重火山群は距離的には近くにあって、緯度や高度はよく似ているが、土地の成因が違うために森林帯は著しい差を生じている。
図3は、大分県の気候を反映した祖母・傾山系の森林帯と、火山作用によって森林帯が変形した九重火山群を比較して画いた大分県の垂直植生帯概念図である。この2つの森林帯を比較することによって、火山作用や人為などによって植生が退行した姿をとらえることができる。
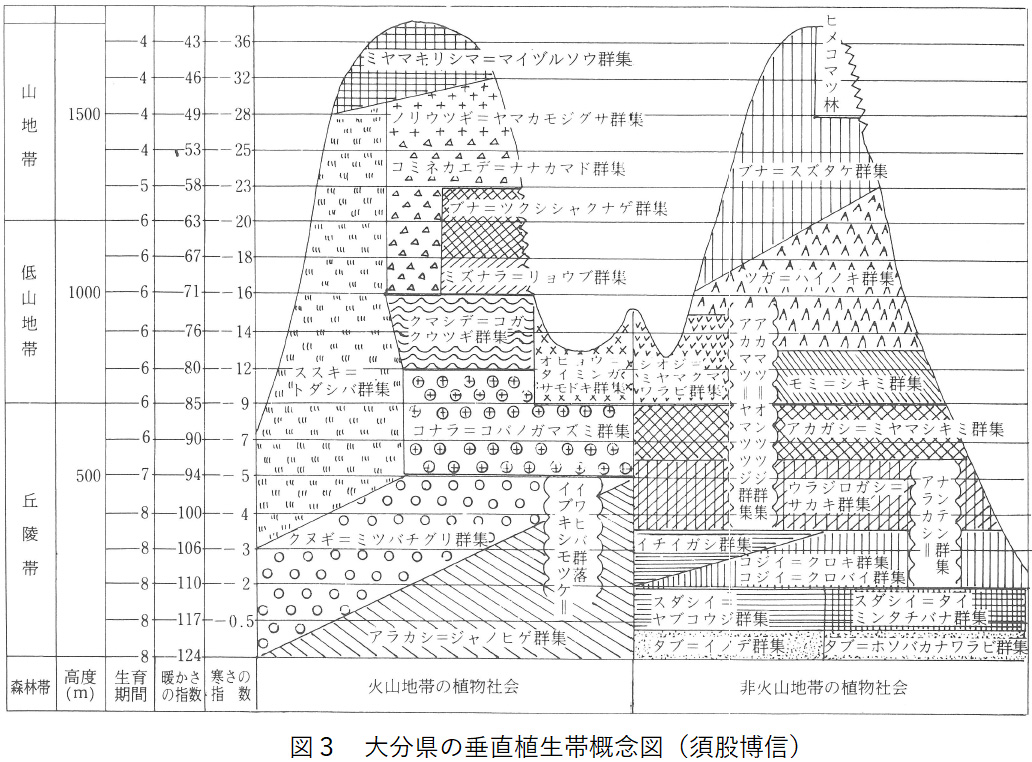
(1)非火山地帯の森林帯
大分気象台の観測資料を基にして、気温減率0.57を用いて大分県規準温度垂直分布表を作り、それぞれの高度における暖かさの指数と寒さの指数を求めて図3に記載した。
月平均気温10℃以上の月の継続をもって生育期間とし、それ以外は休眠期間としてあらわすと、大分県は海面から400mまでは生育期間8ヵ月で常緑広葉樹林の適温である。コジイやスダジイの群落やイチイガシ群集が成立するのはこの温度範囲である。海抜500mで生育期間7カ月となるが、この温度でも常緑広葉樹林は成立する。むしろ空中湿度が増してウラジロガシ-サカキ群集が発達する。海抜600mで生育期間6ヵ月となるが、4月の月平均気温はなお9.9℃であってアカガシ林が成立する。海抜700mでは暖かさの指数85、寒さの指数-9.1になって、この付近が常緑広葉樹林帯の上限になっている。
海抜700mから1200mの間は、生育期間6ヵ月であって、ここではモミ・ツガの針葉樹林が発達する。高木層を形成するモミ・ツガの針葉樹が防寒マントの役目を果たして、亜高木層以下にヤブツバキ、シキミ、ハイノキなどの常緑広葉樹を伴うなど、中間温帯の性格が強い植物社会である。
海抜1000mを越えて休眠期間の月平均気温に氷点下の温度が現れるようになると、常緑広葉樹の生育は無理であって、落葉して休眠するブナが越冬に有利になる。しかし、冬季は晴天が続いて積雪が少なく日照時間が長い太平洋側では、ツガの勢力が非常に強くて海抜1200m付近までツガ-ハイノキ群集が発達する。
ブナは海抜1000m付近から出現するが、発達したブナ-スズタケ群集が成立するのは海抜1200mを越えて生育期間が5ヵ月になってからである。暖かさの指数60、寒さの指数-20付近がツガ帯とブナ帯の境界になっている。
日本のブナ林はカエデの仲間を多く伴い、林床にササ類が多いのが特徴である。積雪は植生を機械的に破壊する場合もあるが、反対に植物を寒波から守る効果がある。雪の下で越冬したササ類は、冷温帯多雨気候に適応したブナと結びついて、日本海側の多雪気候下ではブナ-チシマザサ群集を生じ、積雪量の少ない太平洋側ではブナ-スズタケ群集である。大分県ではスズタケを伴うブナ林が普通であるが、県北部ではスズタケ林床からチシマザサ林床の移行帯に現れるミヤコザサ林床のブナ林も出現する。
日本の亜高山帯を形成するトドマツ、トウヒなどの針葉樹林は暖かさの指数15~45の間、生育期間が1ヵ月以上、4ヵ月未満の高度で発達する。大分県は海抜1700mで暖かさの指数43であるが、海抜1800mになっても生育期間は4ヵ月であって、なお、ブナ林の成立を許す温度条件である。そのため、亜高山帯針葉樹林は成立せず、地形や地質などの条件さえよければ、山頂部までブナ帯になる。
(2)火山地帯の森林帯
図3の左側は、大分県中央部の九重火山群にみられる植生の垂直分布概念図である。火山活動を反映して山頂に近いほど、非火山地帯との植生の違いが大きい。
九重火山群の山頂部は、気候的にはブナ林になってよい高度でありながら、高木が育たないでミヤマキリシマ-マイヅルソウ群集という低木林になる。コケモモ、イワカガミ、マイヅルソウ、マキバエイランタイなどの高山帯植物が生育しており、景観も高山帯の風衝ハイデによく似た植物社会である。これは火山活動と九州最高峰の厳しい気象条件を反映したものであって、鈴木時夫はミヤマキリシマ群団が成立する植生帯を“火山山頂帯”と名付けた。中部日本の高山帯は海抜2500m以上に見られるが、九重火山群では海抜1500m付近が火山の森林限界になっている。
火山森林限界付近には、タンナサワフタギ-ノガリヤス群落やノリウツギ-ヤマカモジグサ群集などの低木林を生じるが、これはブナ林が火山活動や野火などによって退行的に遷移したものであって、火山地帯のブナ帯にモザイク状に出現する。
海抜1000mから1500mのブナ帯にはブナ-スズタケ群集のほかに、ブナ-ツクシシャクナゲ群集を生じるが、これは安山岩が重なってササの繁殖が阻害されるため、ブナとササの結びつきが離れてできた群集である。
若い火山地帯はツガ群団が貧弱なことも大きな特徴の一つであって、ツガ帯からブナ帯の尾根部はコミネカエデ-ナナカマド群集が発達する。また、ツガ-ハイノキ群集やモミ-シキミ群集に代って、クマシデ-コガクウツギ群集やコナラ-コバノガマズミ群集などの落葉広葉樹林が広い範囲にわたって生育しており、常緑広葉樹林帯と落葉広葉樹林帯が接している。
しかし、九重火山地帯にモミ-ツガ林が欠如しているわけではない。九重火山群の北側山麓を流れる鳴子川渓谷側壁では、海抜650~780m付近にモミ-シキミ群集の残存林があり、海抜800~850mにはツガ群落の残存林がある。但し、この生育地は新第三紀中期-第四紀中期の噴出によってできた九重火山基底火山岩類であって、九重火山群本体よりも地質年代が古い。
九重火山群の本体は第四紀更新世後期から完新世にかけて噴出した地質年代の新しい火山であって、有史時代にも噴火の記録があり、今も一部に硫気噴出が見られる。このように若い火山であることと、人類文化の影響を受けて、九重火山群におけるツガ群団の退行現象が起きたと考えられる。
山麓の火山高原は定期的な火入れ、放牧、採草などによってススキ-トダシバ群集が維持されている。また、ススキ草原に隣接してクヌギを植栽しており、クヌギ-ミツバツチグリ群集を生じる。
4.植物社会の解説
(1)九州山頂帯の植物社会
- ミヤマキリシマ-マイヅルソウ群集
- 群集標徴種はミヤマキリシマ、マイヅルソウ、コケモモ、イワカガミ、コイワカンスゲ、ツクシゼリ、へビノネゴザ、カリヤスモドキ、アキノキリンソウ。このうちコケモモ、イワカガミ、マイヅルソウは日本トウヒ-コケモモ・クラスとミヤマキリシマ群団を結ぶ大切な標徴種である。
ミヤマキリシマを上層にもつ低木林で、九州の火山の海抜1200m以高の尾根部や山頂部に発達し、火山山頂帯を形成する。また、九重火山群の山頂部では本州中部以北の高山帯風衝地の代表的な樹枝状地衣であるマキバエイランタイの活力度が高い。 典型亜群集は樹高約1mのミヤマキリシマが樹海状に広がり、イタドリ、イブキトラノオ、イヨフウロ、シモツケソウなど草原要素を伴う。大船山や平治岳の山頂部に発達し群落を形成する。
コケモモ亜群集は、コケモモ、フクオウソウ、ノギラン、コメススキを識別種とする。ミヤマキリシマの樹高は5~10㎝、コケモモと同じ階層で低い起伏の流形状をなして、海抜1500m以上の高度にある尾根や山頂部の風衝地に発達する。相観的には中部高山帯の風衝ハイデによく似ている。典型的な群落は星生山や三俣山頂上部に見られる。また、黒岩山、星生山、肥前ヶ城の尾根部には、優占種にミヤマビャクシン(ツクシビャクシン)をもつミヤマビャクシンファシースを生じる。
ベニドウダン-イワカガミ群落はヤシャブシの勢力が増して、土壌の浅い尾根部に生じる。
(2)山地帯の植物社会
日本のブナ林は、ブナ科とカエデ科の共存とブナとササの結びつきに特徴をもっており、本県ではコハウチワカエデ、スズタケが強く結びついたブナ-スズタケ群集と、土地的条件が悪いためにブナとササの結びつきが離れたブナ-ツクシシャクナゲ群集が認められている。
- ブナ-スズタケ群集
- 標徴種はブナ、コハウチワカエデ、オオカメノキ、ミズナラ、コシアブラ、タンナサワフタギ、スズタケ、シロモジ、コハクウンボク。この群集は太平洋岸気候域における山地帯の冷温帯湿潤気候の極盛相であって、本県では祖母・傾山系、犬ヶ岳、英彦山、御前岳、釈迦ヶ岳、酒呑童子山などの海抜1100m以上の高地にブナ帯を形成する。九重火山群においても存在するが、火山作用や野火などによって退行した代償植生がめだっている。
- ブナ-ツクシシャクナゲ群集
- ブナ-スズタケ群集の標徴種と共通するものが多いが、重要な標徴種であるスズタケを欠いている。独自の標徴種としてはツクシシャクナゲ、ツクシコウモリソウ、フジシダなどがあり、ベニドウダン、コミネカエデ、ナナカマドなどの適合度が高い。
九重火山群の黒岳は自然林に覆われている。この山は火山灰をかぶらないで火山噴出岩上にブナとササの結びつきが離れて、ツクシシャクナゲとブナが結びついてブナ-ツクシシャクナゲ群集が成立する。この群集は九重火山群のほかに英彦山や犬ヶ岳などでも認めらてれいる。 - ミズナラ-リョウブ群集
- ブナ-スズタケ群集標徴種のうち、最も重要なブナ、スズタケを欠くけれども、そのほかの標徴種は共通している。独自の標徴種に乏しいが、リョウブの適合度がやや高く、ミヤマクマザサやミヤコザサと結びつくのが特徴である。ミズナラ-コハウチワカエデ-オオカメノキ-ミヤコザサ群集という階層の結びつきは、この群集の典型林分である。火山地帯でブナ-スズタケ群集が野火や伐採などによって退行した代償植生である。さらに退行が進むとネザサ、チヂミザサ、ナワシロイチゴなどの標徴種をもったタンナサワフタギ-チヂミザサ群集となる。
これらの群集は、九重火山群の海抜800~1400mにかけて成立する。 - コミネカエデ-ナナカマド群集
- 九重火山群のような若い火山では、ツガ群団とブナ群団の垂直植生帯がはっきりしないで、両群団の特徴を少しずつ併せもったコミネカエデ群団を生じる。群団標徴種はノリウツギ、ナナカマド、ベニドウダン。コミネカエデ-ナナカマド群集とノリウツギ-ヤマカモジグサ群集が認められている。
コミネカエデ-ナナカマド群集は、ブナ、スズタケを欠くが、コハウチワカエデ、リョウブ、オオカメノキなどのブナ群団標徴種を伴う。独自の標徴種に乏しいが、コミネカエデ、コシアブラ、ヤマトアオダモ、ナナカマド、ミヤマガマズミ、ヤマアジサイの適合度が高い。九重火山群の海抜1100mから1500mの尾根に成立しており、山頂帯のミヤマキリシマ-マイヅルソウ群集のヤシャブシ亜群集に隣接している。 - ノリウツギ-ヤマカモジグサ群集
- ブナ-スズタケ群集が野火によって退行した二次林である。標徴種はノリウツギ、ヤマカモジグサ、クロヅル、ベニイトスゲ。九重火山群や由布・鶴見火山群の海抜1200mから1500mの山腹や尾根の各所に見られる。定期的な火入れによって生じたススキ群団に隣接することが多い。祖母・傾山系の本谷山や九折越の尾根部では、昔の猟火によってブナ-スズタケ群集が破壊されたあとに成立する。
(3)低山地帯の植物社会
西日本外帯では常緑広葉樹林帯と落葉広葉樹林帯(ブナ帯)の間に、中間温帯の性格をもったモミ・ツガ帯を形成する。降雪が少なく、日照時間の多い太平洋岸の冬の生活には、落葉して休眠するブナよりも常緑針葉樹のモミ・ツガのほうが有利である。好冷湿性の常緑針葉樹と好温湿性の常緑広葉樹が結びついて中間温帯の性格が強いツガ群団が成立する。群団標徴種に乏しいが、ツガ、モミ、ヒメシャラ、アオハダ、アセビ、ツルシキミなどの適合度が高い。
- ツガ-ハイノキ群集
- ツガ、ハイノキ、ヒメシャラ、ソヨゴ、シラキ、アセビを標徴種にもった九州特産の群集である。高木第一層の優占種はツガ、高木第二層はヒメシャラ、アカシデ、イタヤカエデ、コハウチワカエデ、亜高木層はハイノキ、ソヨゴ、シラキ、リョウブ、アセビ、ネジキ、低木層はサイコクミツバツツジ、コバノミツバツツジなどが常在的に生育する。
この群集は祖母・傾山系で特に発達しており、海抜800~1200mにツガ帯を形成する。県北の英彦山、犬ヶ岳、津江山系の釈迦ヶ岳。酒呑童子山、御前岳にも見られたが、国有林の伐採がすすんでわずかに残っているにすぎない。九重火山群や由布・鶴見火山群などのように若い火山では、ツガの生育が悪く、クマシデ、イヌシデ、アカシデ、コナラなどを優占種とする低山地帯の落葉広葉樹林を生じる。また、九重火山群の鳴子川渓谷側壁にツガの残存林が見られるが、ここではツガと常緑広葉樹の結びつきが離れて、ツガ-スゲ型の森林になる。 - モミ-シキミ群集
- 標徴種はモミ、シキミ、アカシデ、イヌツゲ、カヤ。高木第二層や亜高木層はアカシデ、イヌシデ、クマシデ、イロハモミジ、シラキなどの落葉広葉樹のほかに、アカガシ、ウラジロガシ、サカキ、ヤブニッケイ、ヤブツバキなど、スダジイ群団標徴種を常在的に伴う。これらの常在種の有無によって、西南日本外帯的なアカガシ亜群集と、火山作用が強くてやや内陸的なシラキ亜群集に分れる。かつては、祖母・傾山系、御前岳、釈迦ヶ岳、犬ヶ岳、英彦山などにおいて、常緑広葉樹林帯の上部に隣接して海抜650~900mに森林帯を形成していたが、近年は開発がすすんでスギ造林地に変わり、自然林はきわめて少ない。
モミは火山ガスに弱いようで、九重火山群の黒岳や由布・鶴見火山群ではモミを欠く形でこの群集が存在する。しかし、九重火山群の鳴子川渓谷の急崖には、唯一この群集が完全な形で残っている。 - クマシデ-コガクウツギ群集
- 標徴種はクマシデ、イヌシデ、アカシデ、ヤマボウシ、リョウブ、コガクウツギ、ケクロモジ、エゴノキ。由布・鶴見火山群や平家山などの火山灰土の上では、モミ・ツガの生育が悪く、伐採を繰り返すとこの群集に退行する。火山地帯の海抜500mから900mに成立する。
- コナラ-コバノガマズミ群集
- 標徴種はコナラ、コバノガマズミ、クリ。このほかミズキ、リョウブ、ハリギリ、アセビ、ネジキ、コハウチワカエデ、アサガラ、オオモミジ、ウラジロノキの適合度が高くこの群集に結びつく、また、低木層にネザサを伴うことが多い。クマシデ-コガクウツギ群集とはアカシデ、イヌシデ、アワブキ、エゴノキ、ヤマボウシ、ネジキなどが共通している。
- 火山灰地帯の草原が刈草、放牧、火入れをやめるとこの形になり、九重火山群や由布・鶴見火山群の海抜500mから900mに見られる。特に九重火山群の九酔渓や由布山麓の岳本には、コナラの樹高25m、階層構造がよく発達したコナラ-コバノガマズミ群集が成立する。
- スダジイ-タイミンタチバナ群集
- 標徴種はミミズバイ、タイミンタチバナ、ヤマモモ、サカキカズラ。タイミンタチバナはウバメガシ亜群団やコジイ-クロバイ群集にも出現するが、本群集において最高の適合度をもつ。このほかホルトノキ、ホソバカナワラビ、ヤツデ、ノシランも常在度が高い。この群集は伊豆半島以南の太平洋岸に分布し、本県では黒潮の影響が強い佐賀関半島以南の海岸に近い台地に残存林が多い。佐伯市蒲江の王子神社や臼杵市の津久見島はこの群集の範型である。
- スダジイ-ヤブコウジ群集
- 独自の標徴種に乏しいが、スダジイ-タイミンタチバナ群集標徴種の適合度が低く、代表的標徴種のタイミンタチバナを欠如する。組成はコジイ-クロキ群集に似ているが、高木層の優占種がスダジイであることや、コジイ-クロキ群集が内陸部に成立するのに対して、この群集は海岸に近い台地にできて生態的にも異質である。
佐賀関半島以北の海岸に近い台地に分布しており、黒潮の影響が比較的に強く残る国東半島の別府湾沿岸はイズセンリョウ亜群集となる。国東市国東町の桜八幡宮の境内林はその範型である。これに対して国東半島北側の周防灘沿岸では、黒潮の影響は大きく後退してモチノキ亜群集となり、国東市国見町武多都神社に好例を見る。 - コジイ-クロバイ群集
- 標徴種はクロバイ、サザンカ、ツクバネガシ、ヤマビワ。スダジイ-タイミンタチバナ群集標徴種の一部はこの群集に越境してくるが、この群集の標徴種はスダジイ-タイミンタチバナ群集に越境することはないので、はっきり区別できる。生態的にもスダジイ-タイミンタチバナ群集が海岸に近い台地上に分布しているのに対して、本群集は県南部の内陸低地に分布しており、宮崎県境の重岡や宗太郎付近の国有林に広く残っている。また、佐伯市付近や日豊海岸には日本固有種のハナガガシの自生がある。ハナガガシ林は独自の群集を成立させるまでにいたらないで、コジイ-クロバイ群集の1ファシ-スにとどまっている。佐伯市堅田郷八幡宮の境内林はその範型である。
- コジイ-シイモチ群集(シイモチ-シリブカガシ群集の異名群集)
- 標徴種はシイモチ、シリブカガシ、リンボク、オオカグマ、タラヨウ。これらの標徴種はコジイ-クロバイ群集にも生育しているが、反対にコジイ-クロバイ群集の標徴種はこの群集に越境することはないので区別できる。日田・耶馬渓地方や県北の内陸部低地に分布するが県中央部には見られない。
- コジイ-クロキ群集
- クチナシ、モッコクの適合度がやや高いが独自の標徴種に乏しい。上記のコジイ-クロバイ群集やコジイ-シイモチ群集の標徴種を欠落するので区別できる。佐賀関半島の内陸部や別府湾に面した内陸部の台地に残存林が多い。大分市柞原八幡宮の境内林はその範型である。
- イチイガシ群集
- 標徴種はイチイガシ、クスノキ、ミミズバイ、ヤマビワ、センリョウ、ツルコウジ、イズセンリョウ。組成的にはコジイ-クロバイ群集に類似しているが、生態的にはイチイガシ群集が土壌が厚く水分条件に恵まれた場所に成立するのに対してシイ群落は南向き陽光斜面や尾根に生育している。宇佐神宮境内林はイチイガシ群集の範型である。
- アラカシ-ジャノヒゲ群集
- 標徴種は、ナナミノキ、アラカシ、カクレミノ、シュンラン、フユイチゴ、イタチシダ。スダジイ群団の典型亜群団に常在的なスダジイ(またはコジイ)、イスノキ、ツルコウジを欠如して、エノキ、エゴノキ、ヤマハゼなどの二次林要素を伴う。シイ林が成立しにくい阿蘇溶岩台地の急崖や高崎山の安山岩上、あるいは大分川の河岸段丘側壁に発達し群落が残存する。
また、アラカシは若いうちから多くの種子をつくり、萌芽による再生力も強いので、シイ林やイチイガシ林をくり返して伐採するとアラカシ-ジャノヒゲ群集へ退行する。 - アラカシ-ナンテン群集
- 標徴種はナンテン、エノキ、ムクノキ、ビワ、クリハラン、ヒロハヤブソテツ。このほかモモ、ウメ、ユズなど人類文化と関係の深い植物が野生の状態で生育している。臼杵市野津町風連鍾乳洞付近や豊後大野市三重町大白谷などの石灰岩の露頭が見られる場所に成立する。
- ウラジロガシ-サカキ群集
- 標徴種はイヌガヤ、カヤ、ホソバタブ。この群集とアカガシ-ミヤマシキミ群集はウラジロガシ、イヌガシ、バリバリノキ、サカキなどによって、ウラジロガシ亜群団にまとまり、常緑広葉樹林の中では低温に耐え、霧によって日照時間が制限される環境に成立する。また、県南部や国東半島では太平洋型気候を反映してイスノキが優勢でイスノキ亜群集になる。
この群集は海抜350mから550mの丘陵帯谷状地から山腹に生じるが、ここは早くからスギ人工林に開発されたためにウラジロガシ-サカキ群集の残存林は非常に少ない。玖珠町角牟礼城址、由布市庄内町熊群山、大分市九六位山や本宮山、国東半島の文殊仙寺、両子寺、熊野権現など、いずれも神社・寺院の境内林に残っている。 - アカガシ-ミヤマシキミ群集
- 標徴種はハイノキ、ミヤマシキミ(ツルシキミ)、アカガシ、シキミ。ウラジロガシ-サカキ群集よりも高所の山腹から尾根に成立し、モミ-シキミ群集に接して雲霧帯の性格をもつ。豊後大野市緒方町の宮尾国有林、宇佐神宮奥の院の御許山、佐伯市宇目町の鷹鳥屋神社に自然林が残っている。
- タブ-ホソバカナワラビ群集
- 標徴種はフウトウカズラ、ホソバカナワラビ、ホルトノキ、マンリョウ、ヤツデ。佐賀関半島以南の日豊海岸や豊後水道に浮ぶ島に見られる。潮風に対して抵抗力が強い。
- タブ-イノデ群集
- 標徴種はイノデ、クリハラン。高崎山や雨乞岳山麓、別府湾沿岸の豊岡付近の屋敷林に残存林がある。タブノキ群落が成立する場所は海岸に近い低地、あるいは内陸部の山麓扇状地であって、地下から浸出する重力水が森林の生活に利用可能な環境である。
- ウバメガシ-トベラ群集
- 標徴種はウバメガシ、トベラ、ヒトツバ、コヤブラン、シャリンバイ、コバノタツナミ。このほかハマヒサカキ、ナワシログミの適合度が高い。豊後水道域の鶴見半島や四浦半島の急崖では海岸から海抜150m付近までこの群集が占めている。北は佐賀関半島南斜面まで達しているが、それ以北には少なく、杵築市美濃崎に群落の一部が見られる。
- ウバメガシ-コシダ群集
- 標徴種はコシダ、フジツツジ。ネジキやクロマツを常在的に伴ってクロマツ林に隣接する。津久見市や佐伯市米水津で認められている。
ウバメガシ-トベラ群集、ウバメガシ-コシダ群集の両方とも、スダジイ群団標徴種のヤブツバキ、ネズミモチ、ヒサカキ、ヤブコウジ、テイカカズラを伴ってスダジイ群団に属する。
スダジイ群団に属している常緑広葉樹林は暖温帯夏雨型多雨気候に適応した照葉樹林であるが、ウバメガシ亜群団は冬雨型暖温帯乾燥気候に適応した硬葉樹林である。鈴木時夫は「ウバメガシ林が日本列島に来たのは、日本列島が大陸の周辺山脈の陸続きであった第三紀の温暖乾燥の時代であったが、その後、日本海によって大陸から分離して湿潤気候に変ったため、ウバメガシに随伴した温暖乾燥気候に適した植物は滅びて、スダジイ群団の植物を伴うようになった」と考察している。この地域はスダジイ林が優勢であるが、ウバメガシ亜群団が成立する場所は表層土が浅くスダジイの生育は無理である。そのためスダジイ-タイミンタチバナ群集と競合することがなく、豊後水道域のリアス海岸急崖地に遺存することができた。 - ハマビワ-オニヤブソテツ群集
- 標徴種はハマビワ、オニヤブソテツ。そのほかマサキ、ノシランの適合度が高い。スダジイ群団標徴種のヤブツバキ、テイカカズラ、ヤブニッケイ、アオキ、ヤブランを伴ってスダジイ群団に所属する。
佐賀関半島の南岸にはウバメガシ-トベラ群集が分布しているが、北岸にはまったく存在せず、それに代ってこの群集があらわれる。このように本群集は冬季季節風の風上側に発達し、風下側のウバメガシ-トベラ群集とすみわけ的な傾向が認められる。 - シャリンバイ低木林(マサキ-トベラ群集)
- 標徴種はハマカンゾウ、クサスギカズラ、テリハノイバラ、ノジギク。このほかトベラ、ハマヒサカキ、シャリンバイ、ナワシログミ、タマシダ、エビヅルの適合度が高い。日豊海岸の岬や小さな離島の南向斜面は、冬季季節風の風下側による乾燥と、台風などの強風にさらされて高木が育ちにくく、シャリンバイを優占種とする低木林になる。
- シオジ-ミヤマクマワラビ群集
- 標徴種はシオジ、カツラ、サワグルミ、チドリノキ、アブラチャン、ヒメウワバミソウ、ミヤマクマワラビ、ジュウモンジシダ。中央構造線以南の山地と、酒呑童子山、御前岳、釈迦ヶ岳などの津江山地、英彦山、犬ヶ岳方面の海抜700mから1200mの深い渓谷に成立する。近年はスギの人工林に変って自然林は非常に少ない。
- オヒョウ-タイミンガサモドキ群集
- 標徴種はオヒョウ、タイミンガサモドキ(ニシノヤマタイミンガサ)、アサガラ、バイカウツギ。このほかにチドリノキ、ミヤマクマワラビ、ジュウモンジシダを常在的に伴う。九重火山群のように若い火山の渓谷は土壌が移動しやすく、このような場所にあらわれる。九重火山群の黒岳の谷に好林分が残っている。
- トチノキ-ジュウモンジシダ群落
- 九重火山群の鳴子川渓谷では、ケヤキやイタヤカエデと一緒にトチノキが茂る谷型の森林を生じる。組成はシオジ-ミヤマクマワラビ群集やケヤキ-ヒメウワバミソウ群集との類似性が高いが、九重火山群にトチノキを伴う自然林が存在することを強調しておきたい。
- ヒノキ-ツクシシャクナゲ群集
- 標徴種はヒノキ、ナナカマド、ベニドウダン、ネジキ、コツクバネウツギ、アクシバ、シコクママコナ。英彦山や鷹ノ巣山の海抜1000m前後の尾根岩角地形に生じる。ヒノキとツクシシャクナゲの結びつきが強く、ブナ群団からミズナラ、リョウブ、マルバアオダモ、ツガ群団からツガ、ハイノキ、ツヨゴ、イヌツゲなどの標徴種が越境する。表土の浅い岩角地形に残存した第三期遺存植物社会である。
- ヒメコマツ-ヒカゲツツジ群集
- 標徴種はヒメコマツ、ヒカゲツツジ、ツクシアケボノツツジ、ヤマグルマ、ベニドウダン。ブナ群団標徴種のブナ、ミズナラ、オオカメノキ、スズタケ、ツガ群団標徴種のツガ、ソヨゴなどを伴う。祖母・傾山系の海抜1300mから1600mの尾根や山頂の岩角地形にあらわれる。
- アカマツ-オンツツジ群集
- 標徴種はオンツツジ、トサノミツバツツジ、フモトスミレ。アカマツの活力は高く、ネジキ、ウラジロ、カマツカ、サルトリイバラ、コシダ、シャシャンボなどアカマツ群団標徴種を常在的に伴う。亜高木層はヒサカキ、サカキ、コジイ、ウラジロガシ、シキミ、クロキ、ヤブツバキ、サザンカなど、スダジイ群団要素が優勢である。中央構造線外帯の豊後大野市三重町から宮崎県境宗太郎にかけて、丘陵帯の尾根はこの群集が発達している。土壌は浅くしかも菌糸網がよく発達しているため、雨量の多い地域でありながら土壌は乾燥しており、土地的極盛相の性格が強い群集である。
- アカマツ-ヤマツツジ群集
- 標徴種はヤマツツジ、ザイフリボク、ネザサ、コウヤボウキ。このほかアカマツ群団標徴種のアカマツ、ネジキ、カマツカ、アセビ、スダジイ群団標徴種のヒサカキ、ネズミモチ、アラカシなどを常在的に伴う。ヤマハゼ、エゴノキ、ヌルデ、ヤブムラサキなどを伴って二次林の性格が強く、皆伐から萌芽更新するとアラカシ-ジャノヒゲ群集になる。中央構造線内帯のアカマツ林はすべてこの群集であって、夏期の雨量が比較的に少ない国東半島や宇佐地方の丘陵帯尾根では特に発達している。
- イワシデ-ツシママンネングサ群落
- 耶馬渓や国東半島の凝灰角礫岩からなる急斜面は土壌が浅く、乾燥するためにイワシデ低木林を生じる。この低木林にはザイフリボク、マルバアオダモ、ハマクサギ、アラカシ、コツクバネウツギ、ツシママンネングサ、ウンゼンマンネングサなどが結びつく。
- イブキシモツケ-イワヒバ群落
- イブキシモツケ、キハギ、イワヒバ、ホソバヒカゲスゲの優占度が高く、しかも常在的に出現する。このほか低木のマルバアオダモ、ザイフリボク、コツクバネウツギ、オオコマユミなどを伴う。耶馬渓や国東半島の凝灰角礫岩の露岩上に見られる。
- 海岸断崖地の植生
- ボタンボウフウ、ハマボツス、マルバマンネングサ、ハマナデシコなどの標徴種によってボタンボウフウ群団にまとまるが、黒潮の影響を反映して幾つかの群集や群落に分かれる。
ノジギク-アゼトウナ群集はノギジク、アゼトウナの適合度が高く、佐賀関半島以南の日豊海岸全域に広くあらわれる。黒潮の影響がより強い鶴見半島以南ではソナレムグラ、ハチジョウススキ、ヒュウガトウキを識別種にしてソナレムグラ亜群集を生じる。これとは反対に黒潮効果の小さい湾内や入江の断崖地、更に佐賀関半島以北の別府湾や周防灘に面した海岸崖地はアゼトウナを欠如して、ハマカンゾウ、ツワブキなどを共有するノジギク-ツワブキ群落を生じる。また、日豊海岸崖地で海水のしぶきをうけるような岩上のすき間には、イソヤマテンツキ群落が先駆的に生育する。 - 海岸砂地の植生
- 海岸の砂浜は汀線から内陸地に向かって、ツルナ・コウボウムギ群落→ケカモノハシ群落→ハマゴウ群落、その背後はダンチク群落→マサキ-トベラ群集へ続く場合と、クロマツ防風林→ヤブニッケイ林になる場合がある。
海岸の砂浜から砂丘にはハマエンドウ、ハマヒルガオが常在的に生育している。砂の移動が大きい砂浜はコウボウシバ、コウボウムギ、ハマボウフウ、ネコノシタの適合度が高いコウボウムギ-ハマヒルガオ群落が発達し、波によって有機物が寄せられる場所はでツルナ群落を生じる。コウボウムギ群落のハマヒルガオ、ハマエンドウ、ケカモノハシは、ハマゴウ地這性低木林の中でも活力度が高く、チガヤ-ハマゴウ群集の主要な構成要素となって砂丘を発達させ安定化する。これらの植物社会は、別府湾に面した国東半島や日豊海岸の砂浜や砂丘に広く見られる。また日豊海岸の砂丘ではハマオモトを伴ってハマオモト亜群集を生じる。蒲江町のうさかの浜、波当津の砂丘植生はその範型である。 - ダンチク群落
- ダンチク、ハマウド、ハマナタマメ、イタドリ、ツルソバの適合度が高い。ダンチクは別府湾沿岸にもわずかに見られるが、佐賀関半島以南の日豊海岸に発達する。崖下の土壌・砂礫の堆積地や礫浜の背後地などに群生し、マサキ-トベラ群集やハマビワ-オニヤブソテツ群集に隣接する。
- クロマツ林
- 砂防のための人工林であって、クロマツ、トベラ、ナワシログミ、フジナデシコ、カワラサイコ、コマツナギ、ハタガヤの適合度が高い。チガヤ-ハマゴウ群集の内陸側に隣接する。
- 塩湿地の植生
- 波の静かな遠浅の内湾や河口付近の砂泥地は、耐塩性または好塩性の性質をもった植物社会を形成する。
ハマサジ群集は波のおだやかな河口部や砂嘴の内側入江などの塩湿地に見られる。河口部の汽水域ではフクドの優占度が高いが、海岸に近い泥質地ではハママツナ群落、あるいはヒロハマツナ群落になる。また、満潮線付近の有機質に富む場所はホソバハマアカザが優勢であり、冠水する程度の砂泥地はナガミノオニシバ群落が発達する。これらの群落に隣接してヨシ群落が発達するがこのほかにシオクグ群落、シバナ群落、ウラギク群落などが認められている。また、塩湿地の背後地の砂礫上にはアイアシ群落やカモノハシ群落が発達する。 - チゴザサ-マアザミ群団
- ヨシが常在的でチゴザサ、エゾミゾハギ、マアザミ、ヒメシダ、ノハナショウブなどの適合度が高い。この群団は典型亜群団のほかにホソバオグルマ、アカバナ、ヌマクロボスゲなどの適合度が高いホソバオグルマ群落とケナガホノシロワレモコウ、ゴウソ、シロイヌノヒゲなどの適合度が高いナガボノシロワレモコウ群落が認められている。
これらの群落のほかに、ヨシ、ヤマアゼスゲ、ミズオトギリの適合度が高いヨシ-ヤマアゼスゲ群集、及びイトイヌノハナヒゲ、カリマタガヤ、ヤチカワズスゲ、ミズトンボなどの適合度が高いイトイヌノハナヒゲ-カリマタガヤ群集が、火山地帯の海抜600mから850mに見られる。鶴見岳の扇状地である猪の瀬戸湿原、日出生台湿原、地蔵原湿原、久住高原、あるいは飯田高原などに上記の湿原が発達する。また、湯布院町小田の池湿原は海抜770mであるが、泥炭層が発達して、ハリミズゴケ、コアナミズゴケ、ヌマクロボスゲ、ヤチカワズスゲ、ミツガシワなどを伴ったヌマガヤ-ヌマクロボスゲ群集やイヌノハナヒゲ-コアナミズゴケ群集を生じる。 - ヌマガヤ-マアザミ群集
- ヌマガヤ、ヒメミズゴケの適合度が高く、ヨシを常在的に伴う。
九重火山群の坊ガツル湿原に典型的なヌマガヤ湿原が発達しており、飯田高原のタデ原にも見られる。
坊ガツル湿原は、九重火山群に囲まれた海抜1250mの所にある盆地状扇状地である。本群集の典型部はオタカラコウ、マアザミ、エゾミソハギ、ヤマアゼスゲなどの適合度が高いオタカラコウ群落、及びヤチカワズスゲ、サワギキョウ、オオミズゴケの適合度が高いヤチカワズスゲ群落、並びにヌマクロボスゲ群落が優勢である。このほかに、周縁部にシロイヌノヒゲ群落、ヤマアワ群落などが見られる。また坊ガツル湿原周縁部は、踏圧などの人類文化の影響を受けて、ツクシミノボロスゲ群落を生じる。
九重火山群の海抜1600m付近にある西千里浜や白口谷の湧水地は、相観的に北アルプス高山帯に似て、ガキ田の盛衰に伴う湿原植生が見られる。組成はコケモモ、ヒメミズゴケ、カリヤスモドキ、モウセンゴケなどの識別種によって、ヌマガヤ-マアザミ群集のコケモモ亜群集が認められている。また、西千里浜、扇ヶ鼻、白口谷の小沼ではハリコウガイゼキショウ群落を生じる。 - 浮葉植物群落:ヒシ-ガガブタ群集
- 標徴種はヒシ、ガガブタ、ジュンサイ、オニバス、ヒルムシロ。植生は斑点状、または群状であって、同一群落内に浮葉植物と沈水植物を共有する。
- 沈水植物群落:クロモ群落
- クロモ、ホッスモ、キクモ、マツモをそれぞれ優占種とした植物社会があるが、クロモ群落が量的に広く分布している。
- 小型挺水植物群落:アシカキ群落
- アシカキ、ホソバノウナギツカミ、クログワイの適合度が高い。岸辺から水深1m付近までを生育の適地とする。
- 大型挺水植物群落:ヒメガマ群落
- 識別種はヒメガマ1種である。岸辺から水深数10㎝を生育地として、アシカキ群落のフロラを共有する。
マコモ群落も識別種はマコモ1種である。ミズユキノシタ、カンガレイの適合度がやや高くこの群落に結びつき、時には群生する場合もある。 - ススキ-トダシバ群集
- ススキクラス・群団・亜群団標徴種はススキ、ノアザミ、トダシバ、マルバハギ、アキノキリンソウ、リンドウ、サイヨウシャジン、ミツバツチグリ、ネザサ、ワラビ、シバスゲ、ヒメハギなど。群集標徴種はワレモコウ、アソノコギリソウ、フモトスミレ、シラヤマギク、ノギラン。
ススキ-トダシバ群集は定期的な火入れや採草、放牧によって維持されているが、過放牧が続くと組成が変化する。その退行の過程はススキ型草原→ススキ・ネザサ型→ネザサ型→ネザサ・ワラビ型→ワラビ型→スメズノヒエ型となってシバを混生するようになる。 - ススキ-ヒメスゲ群落
- 海抜1300mの坊ガツル湿原の周辺部に生じる。メヒスゲ、ヌマガヤ、コバノギボウシ、ノリウツギの適合度が高く。ツクシゼリ、アセビを常在的に伴う。
- ススキ-ヒロハヤマヨモギ群集
- 由布・鶴見岳の海抜700~1500mに生育している。ヒロハヤマヨモギ、アケボノスミレ、ナガバシュロソウ、キスミレ、エヒメアヤメ、バイカリソウなどの適合度が高い。
- ススキ-ヒキオコシ群集
- 由布・鶴見岳の海抜500~700m、山麓の谷状地に生じる。ヒキオコシ、クララ、クサフジ、ヤブマオなどの適合度が高い。
- 水田の植物社会
- 水田は耕作する夏季から秋季と、休耕する冬季から春季にかけては、環境が著しく異なるために春型植生と夏型植生が認められている。休閑水田に生じる春型植生は、カズノコグサ-スズメノテッポウ群集が代表的である。この群集はスズメノッテポウ、セトガヤ、レンゲソウ、ノミノフスマ、ミノゴメ、コオニタビラコ、キツネノボタン、ヒメジョオン、アメリカフウロ、ムシクサなどが常在しており、更にナズナ、ハコベ、コモチマンネングサ、ヤハズエンドウ、スズメノエンドウなどの識別種をもつ乾燥型植生と、セリ、ウシハコベ、ヒエガエリなどの識別種をもつ湿潤型植生が認められている。
夏季の耕作水田はウリカワ-コナギ群落が代表的であって、ウキクサ、アオウキクサ、ウリカワ、キカシグサ、アゼトウガラシ、コナギ、チョウジタデ、マツバイなどが常在的である。 - 畑地の植物社会
- 田畑や果樹園に共通して生育する植物を前に述べたが、畑地はこのほかにハコベ、エノキグサ、ハキダメギク、カタバミ、ウシハコベ、ハマスゲ、ツメクサ、メヒシバが常在的である。また、春型畑地植生は上記の植生のほかにツユクサ、ホトケノザ、スズメノテッポウ、オランダミミナグサ、スイバ、キュウリグサ、ヤムエグラ、ハルタデなどを常在的に伴い、夏秋型畑地植生はスベリヒユ、コゴメガヤツリ、イヌビユ、エノコログサ、アゼトウガラシ、オヒシバなどが常在的である。
- 路傍の植物社会
- 路傍植生は、エノコログサ、ヨモギ、コセンダングサ、イノコズチ、メヒシバ、オヒシバ、ツユクサ、セイタカアワダチソウの常在度や優占度が高い。空地の植生もよく似ているが、アキメヒシバ、エノコログサ、オヒシバ、メヒシバ、ヒメムカシヨモギなど、一年生植物が優勢である。空き地植生と路傍植生を比較すると、わずかながら路傍植生に多年生植物が増加して、帰化植物が減少する傾向が認められる。
宅地造成地の先駆的植物社会は、ホウキギク、ヤハズソウ、メヒシバ、オオアレチノギクなどの一生年植物群落を形成し、次いでシロツメクサ、ススキ、チガヤ、ギョウギシバ、メドハギなどの多年生植物が侵入して、ススキ草原化がすすむ。
(4)丘陵帯の植物社会
高温多雨の西南日本太平洋岸の気候と夏季乾燥の瀬戸内海型気候は、低地や丘陵地の植生に大きな影響を与えている。また、火山地帯と非火山地帯の植生の対立は高所の植生ばかりでなく、丘陵地や低地の植生にも及んでいる。ウルム氷期に南に追われた常緑広葉樹林は後氷期になって北上したが、火山活動が激しかった九州では植生の再生にさまざまな影響を及ぼした。
県内の低地や丘陵地に残存する常緑広葉樹林はタブノキ、スダジイ、ヤブツバキ、ヒサカキ、ネズミモチ、テイカカズラ、ヤブニッケイ、ヤブコウジ、ベニシダ、ジャノヒゲ、アオキ、モチノキ、カゴノキ、シロダモ、クロキなどが常在し、これらを標徴種とするスダジイ群団にまとまる。
(5)土地的な原因で生じた植物社会
(A)渓谷の森林
(B)岩角地形の森林
(C)海岸の植物社会
大分県の海岸植生は中央構造線の内帯と外帯で著しい差を生じる。特に外帯はリアス海岸で各所に断崖地形を形成し、台風や土用波に洗われて入江には広大な海岸砂地が発達する。また、外洋に面しているため黒潮の影響も大きい。
(D)湿原の植物社会
ミズゴケを伴う湿原は寒冷多湿な北国のものであるが、九重火山のタデ原や坊ガツルのように、海抜1000mを越える湿地にはミズゴケを伴うヌマガヤ湿原がある。本県の湿原を大別すると海抜1000m付近を境にして、これより低地はミズゴケを欠くチゴザサ-マアザミ群団を中核にしたスゲ湿原を生じ、これより高所ではヒメミズゴケが優勢なヌマガヤ-マアザミ群集が発達する。
(E)湖水の植物社会
農業用水としての溜池は、県内各地の人類文化地帯に見られるが、中津・宇佐平野は特に多く、ここには湖水特有の植物社会を生じる。
(F)人類文化によって生じた植物社会
①ススキ草原
県内の山地草原は、県中部火山地帯の海抜600mから1200mの間に広がっている。九重火山群を中心にした久住・飯田地域、万年山を中心とする玖珠地域、由布・鶴見岳山麓の十文字原・塚原地域に発達するススキ草原は定期的な火入れ、採草、放牧などの人為によって形成され、現状が維持されている。
②スギ・ヒノキ林
スギ林やヒノキ林などの人工林は、当初は前植生の名残りの植物が見られるが、スギ、ヒノキの成長とともに組成は単純化して前植生の植物は姿を消し、地域差はほとんどみられない。一般的な傾向として、丘陵帯のスギ・ヒノキ林はシロダモ、ネズミモチ、ヒサカキ、ヤブムラサキ、カナクギノキ、イヌビワ、ハナイカダ、ニワトコ、チャノキ、チヂミザザ、ミズヒキ、ナルコユリ、フモトシダ、ドクダミ、キヅタ、ビナンナカズラなどが常在的である。
③竹林
竹林は竹材を利用するために植栽されたものであって、マダケ、モウソウチク、ハチクが代表種である。竹林に結びつく特有な植物はないが、前植生の常緑広葉樹林の構成種であるヤブツバキ、タブノキ、シロダモ、ネズミモチ、アオキ、ヤブラン、テイカカズラなどスダジイ群団標徴種を伴う場合が多い。また、チャノキ、シュロ、ナンテン、イヌビワ、チヂミザサ、ホシダ、ホウチャクソウ、ヤマシロギクなどの常在度が高い。
④田畑・路傍の植物社会
田畑や果樹園は耕作、除草などによって特異な環境をつくるため、自然植生の侵入を阻むと同時に、ここが生活の適所となって住みつく植物もある。
田畑や果樹園などに共通して生育している植物はヨモギ、ヨメナ、オオイヌノフグリ、オオバコ、ギシギシ、スイバ、チガヤ、オニタビラコ、ヒメジョオン、アレチノギク、アメリカフウロ、シロツメクサ、カニツリグサ、エノコログサ、オヒシバ、メヒシバ、ギョウギシバなどが常在的である。
(日本植生学会会員 須股 博信)
※このページは「レッドデータブックおおいた(2001)」の掲載内容に加筆・修正したものです。